Summary
Echolocation is the process of sending out a high-frequency pulse of sound, which is reflected off solid objects in the environment. By listening for the sound “reflections” it is possible to build up a picture of the environment. In essence, bats use echolocation to “see with sound”, allowing them to navigate through a cluttered landscape (from city to forest) and locate food in absolute darkness.
The details

Echolocation can be broadly described as “seeing with sound”. As early as 1793, Italian researcher Lazzaro Spallanzai demonstrated that, while blinded bats could find their way around their enclosure, deafened bats lost their sense of direction. The term “echolocation” was first coined by the late Harvard zoologist Donald R. Griffin who, back in 1938, used a microphone sensitive to ultrasound to listen to bats. Echolocation is effectively the ability to localize (find) objects based on how they reflect sound. In the case of our visual system, we rely on light reflected from objects around us to see – bats rely on sound reflected from objects around them to “see in the dark”. Bats emit a variety of chirps and squeaks during flight and listen for the echoes. Sound striking close objects will be reflected back sooner and be louder than sound striking a more distant obstacle. Similarly, by listening for changes in the phase of the echo, bats can determine the type of surface from which the sound was bounced back – a hard, continuous object (such as a wall) will produce a sharper echo than softer objects (such as foliage).
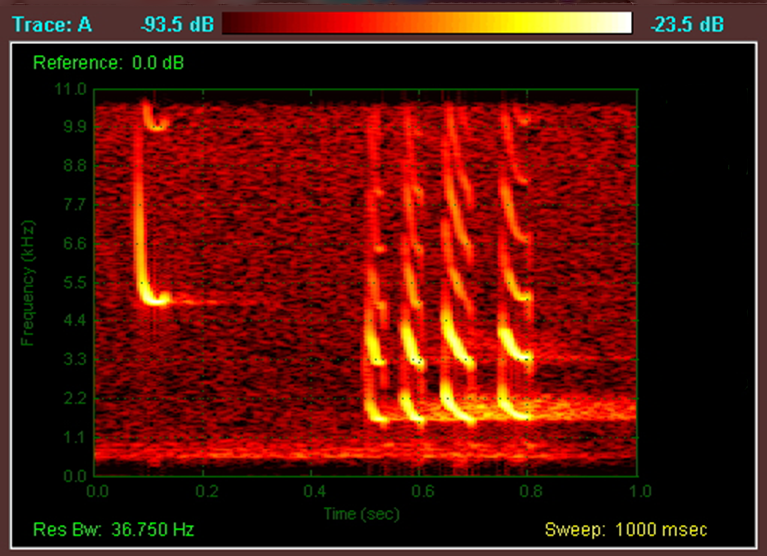
The figure shows the basic principle of echolocation – a sound is produced, bounces back from the first object (the moth in this example) and then, a fraction of a second later, bounces back from a second, third, fourth etc. object (e.g. trees, walls, hedges, etc. in the vicinity). If the bat knows how fast this ‘block’ of sound is travelling, it can calculate -- based on the time separating the two returning echoes -- the distance between the two objects. Moreover, the bat can vary the harmonics, rate, length, intensity and components of the call to gain an extraordinary amount of information about its surroundings.
The frequency -- measured in kilohertz (kHz), or thousands of cycles per second -- of bat calls varies with species and, it is generally considered that, high-frequency sounds give the bat lots of detail but over a short distance, while low-frequency sounds give less detail but over a longer range. Although the rare short-eared trident bat (Cloeotis percivali) of South Africa can call at frequencies as high as 212 kHz -- bearing in mind that humans can only hear sounds as high as about 20 kHz -- frequencies of between 20 and 60 kHz are more common. Frequencies lower than about 20 kHz have a wavelength larger than most insects (so the sound wave moves around the insect, rather than striking it and bouncing back) while frequencies above 60 kHz attenuate (weaken) rapidly in air, which lessens their range.
Bat calls can generally be classed into two groups: narrowband and broadband. Narrowband calls (sometimes referred to as Constant Frequency, or CF, calls) are those of almost constant frequency, while broadband calls (sometimes referred to as Frequency Modulation, or FM, calls) sweep a large range of frequencies in a very short time (i.e. from 100 kHz down to 20 kHz in a couple of seconds). Broadband calls are used to scan the landscape, while narrowband calls are used to identify and provide information on potential prey items. The search calls tend to be intense (with some 10 to 15 calls -- or pulses -- every second in some species), getting faster and faster (up to 200 calls per second) to home in once an insect has been detected.
{kind=link}
In his New Encyclopedia of Mammals, David Macdonald points out that bats tend to have a “Duty Cycle” when echolocating, which represents the proportion of time actually spent generating the sound. In other words, bats only spend a certain period of their time (e.g. about 20%) echolocating, because they can’t listen for returning echoes whilst shouting new pulses. This is not true of all bats; horseshoe bats (Rhinolophidae) can apparently shout and listen at the same time, allowing them to spend as much as half their time echolocating. This is achieved through something known as Doppler Shift Compensation or DSC.
The example I was always given at school to explain the Doppler Shift was the “train approaching a station” illustration – we hear a high-pitched sound as a fast moving train approaches the station platform (because the sound waves are being compressed by the approaching train) and a lower pitched sound as the train races past and away from the station (because the sound waves are being stretched). Thus, the Doppler Shift may be described as the change in pitch (i.e. frequency) of a sound produced by a moving object. How the bats use this DSC is actually rather complicated and involves varying their call frequency according to their flight speed. Sufficed to say that Horseshoe bats send (as ultrasound) and receive (as echoes) sounds on different bandwidths, enabling them to separate their outgoing, echolocative, calls from the returning echoes.
The bat larynx (voice box) is large and reinforced with bone, allowing a high tension on the vocal chords to be maintained (permitting the production of high-frequency vibrations). According to A.A. Wardhaugh’s 1995 book, Bats of the British Isles, sounds are produced in concentrated beams that are directed through a gap in the upper incisors of most species. Horseshoe bats are slightly different – rhinolophids have a flap of skin adorning their nose, which is used to concentrate the stream of sound pulses (in a similar way to a megaphone). This flap of skin seems to afford the Horseshoe bats extra sensitivity, allowing them to detect insects at distances of some 10m (30 ft.), while the vesper (or "evening") bats can only detect insects at distances of about 1m (almost 3.5 ft.). Once the call has been emitted, the ear muscles relax and await the returning echo. Many microbats have a ‘spike’ of cartilage sticking up from the base of their ear, which scientists believe help give the bat better sound detection in a given plane. The echolocation of bats is impressively accurate. An intriguing paper, by Sabine Schmidt at the University of Munich in Germany and two colleagues in October 2000, found that gleaning bats (Megaderma lyra) could find silent and motionless prey on the ground and use their broadband echolocation calls to reject dummy food items whilst hovering over them.
It seems that the ability to echolocate is largely a characteristic of the microbats; megabats (fruit bats, or flying foxes), with few exceptions, don’t echolocate because they have sufficiently good vision to find fruit by sight (scent is probably also involved). One exception to this is the Egyptian fruit bat (Rousettus aegyptiacus), which apparently uses echolocation to find its way about in caves. Indeed, a study by Dean Waters at the University of Leeds and Claudia Vollrath at the University of Freiburg in Germany, found that R. aegyptiacus used echolocation in both light and dark conditions while flying within a tunnel.
Newborn bats appear to pick up echolocation rapidly. In their 2003 paper to the Journal of Neurophysiology, Marianne Vater and five co-workers report that two-week-old moustached bat (Pteronotus parnellii) pups were capable of spontaneously producing CF and FM signals. Dr Vater and her colleagues also report that the ability of these bats to utilize DSC was evident from about four weeks old! It seems that, as well as a rather rapid development of echolocation calls, the call structure can vary according to geography. A 2003 study by Fanni Aspetsberger at the University of Cape Town and two colleagues, found that echolocation calls of little free-tailed bats (Chaerephon pumilus) in the Amani Nature Reserve of Tanzania, were of a lower frequency and had longer gaps between pulses than in those individuals of the same species living in South Africa. These differences are probably related to differences in feeding ecology between the populations.
Bats are not the only animals that use echolocation to find their way about and locate food. Echolocation is perhaps best known in the Odontoceti (toothed whales), especially the Delphinidae (dolphins). In the case of dolphins, sounds (in the form of rapid, high pitched clicks) of about 120 kHz are generated in the nasal sacs, after which the melon (the bony surface of the skull) focuses the sound into a narrow band and projects it forwards. Returning echoes are received by the pan bone of the lower jaw, and are then transmitted to the middle ear by fatty tissue located just behind the jaw; from the ear the sound is transmitted to the brain.
The observation that sound travels four-and-a-half times faster in water than in air suggests that the dolphin’s brain must be extremely well adapted to make sense of the returning echoes, which arrive more rapidly than they do for bats. This may explain why dolphins tend to transmit each click after receiving the echo from the previous one. Some authors have even suggested that the dolphins’ echolocation may have a healing effect on humans. It has been postulated that the ultrasound emitted by dolphins may have a mechanical and/or electro-mechanical effect on the endocrine (hormone) system, positively stimulating it and providing some relief from certain psychological and psychosomatic illnesses. Research into this idea by Karsten Brensing, Katrin Linke and Dietmar Todt at the University of Berlin, however, rejected the idea that dolphins exhibit a behaviour that leads to patients being exposed to ultrasound in doses comparable to those in medical treatments.