Red Fox Territory & Home Range
For any animal trying to eke out a living in the wild there are certain resources that are essential; food, water, and shelter being the main ones. Now, it is highly unlikely that everything that you need will be clumped together just waiting for you to happen across it -- rather, it's almost invariably dispersed across a wide area and may be spread out between several different habitats (forests, farmland, villages, etc.). The area (often, although not always, around a suitable den/nest/shelter site) over which you spend your time wandering, looking for food and water, is loosely termed your home range. In some cases, you might want to secure the resources in this area for yourself (and possibly your family) and this would involve kicking intruders out of all or part(s) of your home range -- these areas are what ecologists call territories.
Thus, despite often being used interchangeably, a territory and a home range are not necessarily the same thing: a home range may overlap with other members of the same species, while a territory generally will not (note that intruders of the opposite sex tend to be tolerated more than same-sex interlopers). In some cases, perhaps in certain seasons, you may choose to defend the whole area against intruders and therefore it and your territory are the same, while at other times (such as when food is abundant) you may allow intruders in some or all areas. In essence, though, the home range is the area transversed by an animal during its normal activity (i.e. finding food, mating and caring for young) and a species is considered territorial if each individual (or group) constructs and defends an area from intrusion by other members of the same species.
Space to roam - territory size

While the size of an animal's territory is largely determined by the distribution of resources across the area, group size is determined by the average abundance of those resources (i.e. how many animals the resources in the territory can support). Consequently, territory size and group size may vary independently and there is no significant relationship between the size of a territory and the number of animals living in it. This is because a small, but rich, territory can support more animals than a larger, but poorer, one. During some seasons or years there may be more resources in the territory than the residents require and this means that other individuals (usually offspring) may be permitted to hang around.
We will come back to this concept in the Behaviour and Sociality section when we look at how group living may have arisen in foxes, but in the meantime I mention it just as a sideline when discussing the flexibility that we see in the territorial system of the Red fox. Indeed, there is tremendous variation in both the territorial behaviour and territory size of foxes; from a complete absence of territoriality in some populations, to complete home range defence in others. Ergo, it is very difficult to come up with the size of an ‘average’ fox territory; if such a concept even exists. That which follows is an overview of territory sizes and "normal" home range movements of foxes -- the subject of how far foxes move when dispersing is covered in a separate section (see Behaviour - Dispersal).
It is now widely accepted that three main factors act in concert to determine the size and shape of the home range of a fox: food abundance; food dispersion; and location of shelters in relation to the main food patches. As a consequence, home range size varies widely: from about four hectares (ha) -- almost 10 acres -- in some urban areas, to 5,000 ha (50 sq-km or about 19 sq-mi) in desert regions, where resources are sparsely distributed. The largest territory of a UK fox that I have found so far is a dog fox in the Eriboll area of northern Scotland who ranged over 4,500 ha (45 sq-km or about 17 sq-mi). In his 1987 book, Running with the Fox, David Macdonald provides some examples of the huge variation in territory sizes:
Boar Hill (gardens and farmland) = 10-70 ha (averaging 40 ha)
Oxford's mixed farmland = 100-250 ha
Wytham Woods (private deciduous woodland) = 20-100 ha (ave. 60 ha)
Oxford city = averaged 90 ha
Suburban Bristol = 25-40 ha (increasing to 90 ha on council estates)
Dutch heathland = averaged 880 ha
Cumbria Fells = averaged more than 1,000 ha
Oman desert = stable range of 5,000 ha
Italian mountain meadows = 400-1,300 ha
North America parks & mountains = typically 600-900 ha
Barren cereal plains of North Dakota = up to 2,000 ha
Arctic = 3,400 ha
So, were we to generalise, we can see that home ranges in Europe typically cover between about 40 ha and 1,300 ha (up to 13 sq-km or 5 sq-mi). This area may contain a single fox, a pair of foxes or, commonly in Europe, small family groups consisting of three to six related animals. Territories tend to be stable during the spring and summer, coming under increasing pressure during the autumn and winter as dispersal of cubs and long-range movements (in search of mates) increase; a difference less apparent among group-living foxes. Where foxes live singly, males seem to range over larger areas than females. In Edinburgh during the early 1980s, for example, Hugh Kolb found that average dog fox territories were 460 ha (just over 4.5 sq-km or almost 2 sq-mi), while vixens ranged over only about 150 ha (1.5 sq-km or just over half sq-mi). A study tracking foxes in the Netherlands found that males were significantly more territorially active than females. In contrast, however, a tracking study of foxes between May and September 1989 in central Kyushu in southern Japan found no significant differences between the territories of dogs and vixens.
Shoo or share? Territory exclusivity
The question of how exclusive territories are (i.e. whether all, part or none of them are actively defended) is deceptively complex and the tremendous flexibility in fox society means there is no "one size fits all" answer. In some cases, such as those encountered by David Macdonald while tracking foxes in urban Oxford, they never seem to trespass -- in his Running with the Fox, Macdonald noted that neighbouring groups didn't trespass, even where some groups had food put out in feeding stations that the others must have been able to smell. Thus, it would seem that, if you have several foxes visiting your garden at once, they're likely to be members of the same social group.
Macdonald considered that habitat type was an important factor in determining territory exclusivity and, in his book, he described how territory boundaries may be "hazy" in rural (especially woodland) habitats, with neighbouring groups using the same clumps of trees and having paths that criss-cross. In urban areas, by contrast, streets provide clear demarcations of territories. He concluded that there was no trespassing in urban areas (where high-quality feeding sites were at stake, territory boundaries were sharply defined and meticulously observed), while deep in the woods (used mainly for sleeping by day) there was "more latitude for neighbourly tolerance". This would certainly explain why, while tracking vixens in the Welsh mountains, Macdonald found that two neighbours occasionally used the same resting sites, although not at the same time. Studies on urban foxes elsewhere, however, paint a different picture.

During 1978-79, for example, Stephen Harris tracked seven breeding vixens in Bristol, finding that they moved, on occasion, just over 300 metres (990 ft) outside their normal ranges and shared feeding sites with animals from adjacent groups. Harris concluded that, probably because the diverse pattern of food availability in urban areas is difficult to defend, the breeding vixens exhibited little, if any, territorial behaviour. Overall, the broad picture seems to be that foxes living in small ranges (at high densities) are more likely to maintain rigid territory boundaries than those living in larger ranges (at lower densities), although there is considerable variation between populations. Off-hand, it would seem logical that smaller areas are easier to defend than larger ones and this may be part of the explanation.
Influences on territory configuration
A territory can be large or small and may or may not be rigorously defended, but what determines its size and shape? We have seen that habitat plays a crucial role in terms of both landmarks to use as borders and the availability of food, but there is another important factor only recently alluded to: the size of the animal. In a 2008 paper to the Journal of Mammalogy, a team of Bristol University biologists led by Graziella Iossa described how: "Increased body size in males appeared to confer an advantage in territory acquisition and defense ...". In essence, the biologists found that smaller males suffered more aggressive challenges for space (what the authors termed "boundary pressure") than larger males and, as a consequence, tended to hold smaller territories. So, larger males held larger territories and made it more difficult for smaller males to secure space.
Earlier work on the take-over of fox territories conducted by Eric Preston in a large fenced enclosure in North Dakota during the summer of 1972, found that it was almost invariably the resident males that tackled intruders and, rather than expulsion being rapid and violent, it was: "...a rather gradual exclusion resulting from continual harassment by the resident male". Preston's findings are at odds with many of the observations that David Macdonald and Stephen Harris have made (both documenting rapid, violent expulsion of intruders -- see Behaviour), but it does give an indication of how the boundary pressure from larger males described by Iossa and her team might operate.
Given that territory size, shape and level of defence are dependent upon the availability and distribution of resources, it is not unexpected to find that foxes living in the food-rich habitat of the city tend to forage over smaller areas -- about 50 ha (half a sq-km or one-quarter of a square-mile) -- than their rural counterparts. There is, however, considerable variation in urban territory size and, in their 2010 overview of fox biology in Urban Carnivores, the Bristol University biologists give the following as territory sizes for urban foxes: 18-169 ha (Bristol); 54-93 ha (Oxford); 259 ha (Virginia, USA); 379-547 ha (Illinois, USA); and 420 ha (California, USA).

Urban territories are generally small with little or no overlap but, as Harris has alluded to above, there are areas (those with abundant food) where animals from neighbouring groups may come to feed without any apparent hostility. In some cases, where food is particularly abundant, so many foxes may be drawn in that it becomes impractical to defend the area and the only option is either to leave or tolerate the intruders. Another result of high food availability and small territories in urban areas is that fox densities tend to be higher; more foxes mean more competition for space and, in a highly tumultuous environment such as the city, where lots of vehicle activity means foxes generally live for less than two years, we sometimes see an interesting feature known as territorial drifting.
Radio-tracking studies of foxes in Oxford city by David Macdonald and Patrick Doncaster during the late 1980s and early 1990s demonstrated that territories were arranged in a "honeycomb" pattern and, although territories in the suburbs appeared spatially stable, the location of those in the city centre continually drifted. Moreover, the biologists discovered that the territories drifted in synchrony with each other (at a rate of about one territory per year) and, although the territories were reasonably small (averaging only 39 hectares -- just under half sq-km or one-fifth sq-mi) they were constant in size. Thus, in Oxford city at least, it seems that as one part of the territory is yielded by fox "A", this area is taken over by one of its neighbours (fox "B"); but fox B doesn't range over a larger area, rather it relinquishes a similar sized area of its own territory. Doncaster and Macdonald suggest that:
"the ultimate explanation for drifting is that the city environment is inherently disturbed, by pedestrian and road traffic, habitat management and construction and demolition work."

It is an interesting thought that such drifting may not be solely the result of foxes dying, but may also be affected by their changing landscape. Indeed, given that conspicuous linear features (e.g. roads, hedges, walls, etc.) are often used as territory boundaries, it is easy to see how the continual state of development in our major cities could lead to the continual fluctuation of fox territories. It seems that there may also be some seasonality to this drifting and, in his 1996 review of the Red fox social system, Paolo Cavallini noted that sudden home range shifts were more common during June to December, while drifting was faster in winter. The picture in Oxford, however, doesn't appear to be universal and, in the West Midlands, the exact opposite has been documented; with territories remaining stable for many years.
In Bristol, the arrival of sarcoptic mange offered an opportunity to see how the fox population responded to a drastic decline in numbers. Prior to the appearance of mange, territories of Bristol's urban foxes were found to be stable across years, with little overlap. In early 1994, a young male infected with mange arrived in Bristol, having spent the winter outside of the city; the disease spread rapidly through the population, causing the death of some 95% of the resident foxes. Unlike the foxes Macdonald and Doncaster tracked in Oxford, however, Bristol's foxes responded to the death of a neighbour by expanding their territory; essentially taking up the slack.
The biologists at Bristol University's Mammal Research Unit observed that, despite the increased level of food availability, foxes continued to expand their territories as neighbouring groups died; survivors took over this free space within a matter of days, enlarging their own territories until they came across an occupied territory. So, why would the foxes do this? They didn't need the extra food (after all, their own territories before the die-off supplied them with more than enough) and a larger area is more difficult to defend, requiring more energy to patrol. The Bristol biologists suggest that the foxes may have been attempting to prevent new animals from settling in the area by monopolizing space. In addition, in the pre-mange period, some foxes were observed splitting their territories -- half being "given" to the resident's offspring -- and if you hold a larger area, any future splitting is simpler and less costly in terms of resource loss.
So, fox territories may be very stable from year-to-year, they may drift as the habitat changes or they may expand as neighbours die. Additionally, some studies suggest that if a territory becomes vacant, and isn't amalgamated into a neighbour's range, the space is quickly filled by a new individual -- studies by Andrew Wilson suggest that this happens within about a fortnight. How, then, do foxes stake out their territory? How does one territory holder know where their turf stops and their neighbour's starts? How do residents let passers-by know this spot is occupied?
The smell of home - scent marking territories
We have already seen that foxes often use linear landscape features such as hedges, tree-lines, roads and so forth as territory boundaries, but the territory owner still needs to let others know this is their hedge and what happens when there's no convenient feature? Given that permanent monitoring of territory borders isn't an option (foxes have much more pressing demands on their time), a convenient signpost is required in the owner's absence; for this foxes use scent to signify occupancy, leaving scent marks in the form of urine and scat at strategic sites throughout their territory. I don't plan to delve too deeply into this topic here (see QA in preparation), but essentially foxes leave scent marks on travel routes in proportion to how frequently they're used and where in the social order the animal sits (dominant animals scent mark more than subordinates).

Droppings and urine are most commonly used as territory markers. An important feature of scent marking is that it should be obvious to others and this explains why most are associated with structural characteristics in the environment, such as landmarks. Indeed, droppings are almost invariably -- about 80% of the time in some studies -- left on visually conspicuous objects such as rocks, tufts of grass, garden gnomes, etc. Urine may be deposited on conspicuous objects, or squirted into vegetation at about nose-height. Droppings make more obvious and more robust signals than urine and so the two excretions may be used differently. Fox urine, scat and the secretions from various glands, including paired anal glands that can excrete on scats that the fox wants to use as territorial markers, contain hundreds of chemical aromas that convey a wealth of information about the individual who left it, as well as how long ago it was made. We know, based on work by Janosch Arnold for his Ph.D. at Bristol University, that fox scent marks contain information about the sex, season, relatedness, health and probably the social status of the animal who left it. It has also been suggested that resident foxes may recognise territory boundaries by the presence of "alien" urine.
Working with his captive vixen, David Macdonald found that the frequency of scent marking fell as you approached the border of her territory (where she would pause for a few seconds before spontaneously turning back). Macdonald described how the exact turning point could be changed by introducing a sample of alien fox urine near the border. If the alien urine was introduced just inside the boundary of her territory, she'd turn back when she smelled it (but would ignore her own transplanted urine), even if the area further on had previously been part of her territory. There was a limit, however, and if the alien sample was introduced elsewhere in her territory -- i.e. not near the border -- it was over-marked (urinated on top of) by the vixen. So, it seems that foxes scent mark most in the core of their territory -- where they're most active -- and less frequently at the periphery, with foxes having a fairly good idea where their territory ends based on where they expect to encounter foreign urine.
Scent isn't, of course, a permanent feature in the environment and after a while it gets degraded by rain, wind, etc. Consequently, the owner of the territory must revisit territory boundaries frequently to reapply the scent - at the same time, failing to do so will result in a weaker scent and might suggest to an intruding animal that the owner is either no longer around, or rarely visits this particular part of their range. Indeed, Macdonald, in a 1980 paper to the Symposia of the Zoological Society of London, suggested that the strategic placement of these scent marks may allow an intruder to chart a "safe" route through a territory by "reading" fresh urine marks; thus avoiding the resident. Indeed, data from field studies have shown that scent marks aren't deposited at random throughout the territory.
Between October 2005 and April 2006, Raquel Monclús at the Universidad Autónoma de Madrid and her colleagues looked at the scent-marking behaviour of foxes around rabbit warrens in central Spain and found significantly more fox faeces where the density of rabbits was higher. Foxes associated their scent marks to rabbit remains, paw scrapes and latrines significantly more than expected and more often than would be expected if at random. In their 2009 paper to the Journal of Ethology, Monclús and her co-workers suggest that foxes visit the rabbit warrens and scent-mark them more than other areas of the habitat and they can then use this high-density scent-marking to keep track of where the prey is within their territory and configure boundaries accordingly. Essentially, that the foxes were using the warrens as amplifiers for their scent marks. The researchers also observed that foxes marked more frequently in clearings than on roads. Clearings are considered strategic marking sites because they represent the intersection of multiple small tracks out of peripheral woodland.
Neighbour recognition?
Generally speaking, neighbouring territory holders have little to gain from fighting with each other (each territory has sufficient resources to support the animal and the potential costs of fighting can be huge) so once the initial battle has taken place to establish where the boundary should be, scent marking offers a conflict-free method of letting each other know that you're still around. Less time spent fighting also means that more energy can be devoted to finding food.

In a paper to the Canadian Journal of Zoology during 2011, Janosch Arnold, Carl Soulsbury and Stephen Harris report that foxes change their behaviour when unfamiliar urine is encountered in their territory. The researchers applied artificial fox urine to known fox territories to simulate the presence of an intruder and radio-tracked the residents to see what they did. The dog foxes didn't change the size of their territory, or the amount of time they spent active. They did, however, shift their daily activity range so they spent more time in the area where the fake scent had been applied; this was more pronounced in big males than smaller ones. The foxes searched a greater proportion of their territories on nights after the scent was applied than before the scent arrived -- presumably looking for the intruder. Vixens showed no significant change in either their searching behaviour or activity range, suggesting that territorial defence is more male-centric.
Territory borders may be recognised by the presence of alien urine or, perhaps equally as probable, the presence of a familiar urine that the resident associates with a neighbour. In a recent note to BBC Wildlife Magazine, Stephen Harris noted how, when the foxes and badgers first appear in his garden in the summer, they sniff each other carefully and then ignore one another, but get very agitated when a stranger turns up. This, and many similar observations from other researchers, suggests that foxes are capable of recognising familiar and unfamiliar animals and that neighbouring territory holders may thus be able to recognise each other. Indeed, during the mid-1970s University of Washington psychologist David Barash trapped seven foxes and held them in cages so he could observe their reaction to each other. Barash found that foxes trapped near each other (within 8 km / 5.5 mi) showed dominance-submissive behaviour towards each other, while those trapped further apart showed more aggressive behaviour (threat gapes) to each other.
Despite the small sample size, Barash's data suggest that foxes can recognise their neighbours and that there may even be a rudimentary social structure between them. This is perhaps not surprising as, in urban areas at least, neighbouring groups are often related. Whether or not an intruding fox recognises the resident, several authors have described how invading individuals change their behaviour when they enter another fox's territory, moving more erratically and ceasing all scent marking. In 1972, the late-great zoologist Niko Tinbergen described how territory holding foxes on the sand dunes of the Ravenglass-Drigg Sanctuary in Cumberland knew their ranges very well and always used the same tracks (often in almost straight lines across open sand) from their earths to favoured hunting grounds; he sometimes found what amounted to "fox highways" along the shoreline. Newcomers to the area, Tinbergen noticed, acted quite differently:
"A fox entering a strange hunting ground changed its behaviour: instead of running in a straight line across the open sands it would try to keep under cover and would move hesitatingly and follow an irregular route."

Interestingly, there are also observations of foxes urinating in such a way as to suggest they may be trying to cover up their scent.
Adele Brand in Surrey and Catherine North in Manchester captured well-grown fox cubs urinating in a water bowl on their trailcams during August 2018 and 2019, respectively. Similarly, West Sussex-based fox watcher Onyx Stewart tells me one of his adult vixens consistently did this in a tub of water in their garden, and Wildlife Online reader Tom Fitton has seen foxes do this during the summer in his garden in London. There was also a curious report of a fox approaching, and then urinating in, a cup of tea left on a kerbside in Wythenshawe while the owner worked on his car during December 2019. Along what I take to be similar lines, I have also received a description of "a nervous young fox" urinating into a drain in Hampshire a few years ago. The fox was seen from a flat window and the observer didn't investigate the drain afterwards, so cannot be certain the urine entered the drain rather than being deposited on the grate, although this was the opinion of the witness. More recently, in September 2023, Adam Taylor watched a vixen pull off a small drain cover and urinate into the drain in his London garden.
Given that foxes mark food remains and conspicuous objects within their territory, this behaviour may be interpreted as territorial marking intended for any other visiting foxes; drain covers, cups and water bowls are prominent objects within a territory, the latter likely to be visited particularly regularly during the summer when this behaviour appears most pronounced. Similarly, foxes meticulously keep track of their territory configuration and doubtless any new objects are marked to help the fox familiarise itself with them. The latter likely accounts for the urinating in the cup of tea. At this point, I cannot explain urination into a drain, particularly going to the trouble of removing the grate first.
Accepting the foregoing, however, if the urine mark is intended as a message, why dilute it? Granted foxes have a remarkable sense of smell, plus a well-developed vomeronasal organ, and are likely to detect the urine even in a bowl of water, but being mixed with the water must surely reduce its volatility and potency compared with, for example, urinating next to the bowl, which would presumably be detected by any other foxes visiting the resource. Furthermore, Tom Fitton tells me that other foxes (and badgers) drink from the bowl despite the contamination, without apparently over-marking as they would ground-based scent marks, perhaps suggesting the scent is much less perceptible. Recently, Kathryn Sobieski got in touch to describe how foxes visiting her property urinate and defaecate on the edges of food and water bowls, some of which inevitably enters the bowl itself, and this does not put foxes off using them. Similarly, one might assume urine ejected into a storm drain is less likely to be detectable to other passing foxes, although Dr Sobieski also referred to an Alsatian she used to have who urinated down drains in what she interpreted to be an innate reaction to cover up the smell of sewage.
It might be the case that foxes aren't aware that urinating into a water bowl or down a drain obscures their scent, but marking behaviour appears an innate behaviour (i.e., not one cubs learn from their parents) and yet "water-urination" seems comparatively rare, particularly among adults. Indeed, the observation that it tends to be subadult animals (i.e., those under a year old) exhibiting this behaviour during late summer raises the intriguing suggestion that it might be a way to "hide" or "dilute" their scent as they reach maturity on their parents' territory, or while trespassing looking for their own territory, thereby reducing the potential for conflict. I would be interested to hear from readers with experience of this behaviour, particularly by adults during the winter or spring.
Not only do foxes know what area their own territory covers and when they're trespassing in someone else's domain, there are some data suggesting they know where their territories are even if they're picked up and dumped somewhere else -- in other words, that foxes have a homing ability. In Minnesota, USA, for example, wildlife biologists Robert Philips and David Mech found that a vixen they captured, translocated and released in November 1968 travelled the 56 km (35 miles) back to her original capture site within 12 days. Similarly, in his 1906 book Foxes at Home, Colonel J.S. Talbot told of a vixen and her cubs that returned to their earth "in due course", having been trapped and relocated eight miles (13km) away. Quite how these vixens knew where their territories were remains a mystery, although it has been shown that European badgers (Meles meles) have a good idea where their territory is relative to those of their neighbours, and can use this knowledge to find their way home; foxes may retain the same information.
Seasonal variation in territory use
So we've seen how big a territory can be and how foxes mark it out, but how much of its territory does a fox use? This, like everything else we've tackled in this section, is not an easy question to answer either. There are many factors that influence how much of a territory gets used and how far a fox will travel each night, although the impact of season is perhaps most significant. The vast majority of tracking studies show the same trend: during the breeding season (winter) foxes travel further than they do at other times of the year. Indeed, during the breeding season males are known to move over large areas -- regularly trespassing -- as they search for receptive vixens, while females typically spend a greater proportion of the time at the periphery of their territories (again, this probably facilitates mate-finding).

Studies in North America have found dogs increasing the length of their nightly movements by four to eight times come the breeding season. Similarly, tracking of Bristol's foxes has found that average winter ranges of males were more than twice the size of those in spring, summer and autumn (although caution should be used owing to low sample size) as they trespassed into neighbouring ranges in search of receptive females with which to mate -- there was no significant difference in the seasonal range of females. Interestingly, although no seasonal difference was found for females in Bristol, this may indicate that none became pregnant during the study. Several studies have demonstrated how pregnant foxes move shorter distances around the time they give birth and for several days after the cubs are born.
Working with the foxes of the Ashio Mountains in Central Japan, for example, Masahiko Takeuchi and Masaaki Koganezawa found that their pregnant vixen's range had decreased by six-fold by the time her cubs were about to be born (pre-breeding range was 601 ha, declining with the advance of her pregnancy to 109 ha by the time her cubs were born).
Seasonal variations in ranging may, however, be for reasons more subtle than searching for a mate or the birth of cubs. Studies in Europe have shown that, during the late summer and autumn, foxes feed heavily on fruits that are locally clumped (they can strip a bramble bush of blackberries in no time); the result is that they need to move around less than they do during the summer or winter (when finding food for cubs and when food is more difficult to find, respectively).
Between 1992 and 1994, Hideharu Tsukada at Hokkaido University studied the interaction and territorial behaviour of 42 foxes in Japan's Shiretoko National Park. In his 1997 paper to the Journal of Ethology, he described a flexible territorial system associated with the distribution of food. When food was not concentrated, resident foxes were highly territorial, showing exclusive distribution of home ranges between families and defence against other foxes at the boundaries. When food was concentrated, such as during the salmon spawning period or when tourists were feeding foxes in the park, home ranges overlapped and foxes made round trips of up to 8km (5 miles), lasting up to three days, from their territories to take advantage of the localised concentration of food.
Interestingly, when one male fox died, his territory was absorbed by neighbouring animals after about eight days. This perhaps explains why Tsukada observed that most foxes making trips away from their range to use seasonally concentrated food sources were away for only a day on average; three days maximum. Food sites more than 8km away weren't utilised and, in the paper, Tsukada suggests:
"This suggests that available feeding sites for foxes were within the area that foxes could reach in a day. This suggested that localized food in other families' home ranges could be utilized by foxes without a high risk of losing their territories."
These paws are made for walking
Generally speaking, foxes move roughly the same distance each night, although adverse weather conditions (especially snow cover) may reduce the distance travelled. In Bristol, foxes moved an average of eight kilometres (5 mi.) per night, rarely exceeding 10 km (6 mi.). Similar values have been recorded in Japan (6-8 km) and Spain (4-5.5 km / up to 4 mi.), with slightly longer trips (up to 15 km / 9 mi.) in more remote European and North American environments.

In the UK, the longest distance travelled by a fox that I have come across in the literature was a male that was tagged as a cub by Huw Lloyd at an earth in mid-Wales and was later shot by a gamekeeper 52 km (32 miles) to the north-east. More recently, however, Brighton University's Dawn Scott tagged an adult male fox in suburban Brighton that was tracked a staggering 315 km (195 miles) from Brighton to Rye in East Sussex in just under one month. This particular fox, known to BBC Winterwatch viewers as Fleet, was believed to have been ousted from his territory by his son (possibly as a result of his suspected lungworm infection); he left Brighton on 9th December 2013 and in two days had reached the rural setting of Ditchling Beacon on the South Downs, some 7 km (just over 4 miles) away.
Fleet spent a couple of days on the South Downs Way before heading north-east, through various towns and villages and, by 18th December, he had travelled some 200 km (124 miles) from his previous territory. He moved south briefly, towards the A27, and then back north to spend Christmas Eve and Christmas Day at a caravan park near Uckfield, where he was presumably fed by, or scavenging food from, residents. The track continued to just north of Westfield in East Sussex, where the signal was lost during early January 2014; a total distance of 315 km (196 miles) and by far the longest verified dispersal by a single fox in Britain. Moreover, Fleet's impressive ramble tops the 203 km (126 miles) by a vixen in North Dakota, of the longest dispersal distance recorded by Stephen Allen and Alan Sargeant, and is not far off that of a dog fox tagged in Wisconsin and recaptured nine months later 394 km (246 miles) in Indiana. It should be noted that, despite a fairly extensive search of the area where the final signal was received, there was no sign of Fleet's body, suggesting that the collar may have failed, and Fleet's journey may still be in progress. It may also be that there was some human interference (e.g., someone shot the fox and move the carcass), but there was nothing about the track of the GPS to suggest this.
More recently, between 2011 and 2017, Zea Walton and her colleagues at Inland Norway University of Applied Sciences tagged 101 foxes of both sexes within four areas of Norway and Sweden with GPS transmitters. Their data, published in the European Journal of Wildlife Management in 2018, show that cumulative dispersal distances ranged from 132 to 1,036 km (82-644 miles), with straight line distances of between 67 and 294 km (42-183 miles) during dispersal events ranging from seven to 22 days. Two foxes subsequently doubled back to settle in an area they had previously explored. Interestingly, while these tracking data highlight the mobility and dispersal capability of foxes across large scales, genetic analysis on the same population by Walton and her co-workers also revealed low levels of genetic differentiation, implying that relatedness plays a strong role in the spatial organisation on a local scale. This was particularly so for females, the researchers finding male-biased dispersal in this population.

Foxes may temporarily leave their territory if there are highly concentrated food resources nearby. Foxes in Japan's Shiretoko National Park, for example, were observed to leave their territories for up to three days while they foraged on concentrated food sources (salmon carcasses and human food) elsewhere in the park, although they didn't stray more than eight kilometres outside of their range. Overall, it seems that foxes focus most of their time at specific parts of their range (generally feeding and resting spots), so they may move several kilometres, but stay in a relatively small area, often using only part of their range.
Nightly patrols or flying visits?
When foxes do move around during their nightly activities they do so along a series of well-defined paths that are about 30 cm (12 in.) in diameter and on which the grass has a "trodden down" appearance. Commonly foxes will use the same paths on successive nights if they lead to a favoured hunting spot, although my experience is that they are less predictable in their habits than badgers. Foxes often opt for the path of least resistance while commuting, using field margins, badger or deer runs, or the "tramlines" of vehicles that have driven across the field.
It is sometimes assumed that, as part of its nightly wanders, a fox will travel in a circuit around the territory, essentially patrolling the borders. In some cases (where territories are small and the fox density is high) this invariably does happen, but many studies have found that foxes focus the majority of their activity in a core area (a breeding earth or rich hunting ground, for example). Studies both here and in the USA have found that, in most cases, foxes don't visit their territory borders every night; in a large territory the fox may only visit a border area once every few days, although the movements of adult members of a group appear to collectively result in visits to all borders every week-or-two.
Even in small ranges, the fox invariably doesn't use the entire range every night. Indeed, in 2002, Polish biologist Jacek Goszczyñski reviewed the published data on fox home range and found that foxes living in small ranges (up to 150 ha) used 36-70% of the range daily, while those living on larger ranges travelled over similar sized areas but that these represented a much smaller proportion of the seasonal range (only 16% of the largest home range included in the analysis was used on any single day). So, the foxes moved over roughly the same sized area each night, regardless of how large their territory was, even if that meant some areas were almost never visited.
Non-territorial foxes

It should be noted that, although territoriality is well known in Red foxes throughout most of their range, it is not observed in every population. Between January 1989 and April 1992 (with an intermission during the Gulf War), a team of biologists from Oxford's Wildlife Conservation Research Unit led by David Macdonald conducted a detailed study on the Red foxes of the biological reserve of Thumamah in the great sand deserts of Saudi Arabia (Vulpes vulpes arabica).
Over a period of three years, 41 foxes (21 males and 20 females) were fitted with radio collars and their movements tracked, revealing an average home range of 676 ha (almost 7 sq-km or just over 2.5 sq-mi) that didn't vary according to sex or season. The most interesting discovery was the lack of territoriality; instead they observed loose-knit groupings, with females sharing territory more readily than males. The biologists suggested that this situation arose because it helped the foxes, which are smaller than Red foxes found in Europe (i.e. about half the size of those in the UK), survive in an extremely harsh environment with a spatially and temporally sparse food supply.
Finally, it is worth mentioning that, even where territoriality is commonplace, not all foxes partake. Indeed, it is estimated that some 15% of adult Red foxes wander with no fixed home range; these animals are called itinerants. Little is known about the behaviour of itinerant foxes, although studies by Andrew Wilson during the mid-1990s suggest that these animals behave differently to territory holders in their habitat use.
In his bio of the Red fox in the RSPCA's 1981 British Mammals book, David Macdonald notes that itinerants are usually males and may move in a straight line or weave a meandering course through the countryside, ranging up to 250 km (170 mi.); he gave average ranges of 20–50 km (13.5–34 mi.) for males and 7-15 km (5–11.5 mi.) for females. Macdonald does point out that the distinction between itinerants and residents is blurred, with some territory holders commuting long distances between centres of activity. Indeed, in a contribution to a 1980 special edition of the journal Biogeographica on Red fox ecology, Huw Gwyn Lloyd noted that, although it's often perceived that foxes are either sedentary or itinerant, in reality an individual can be intermittently itinerant; in mid-Wales he tracked a dog fox making frequent excursions out of his territory (up to 16 km away), but always returning to his familiar range sooner or later.
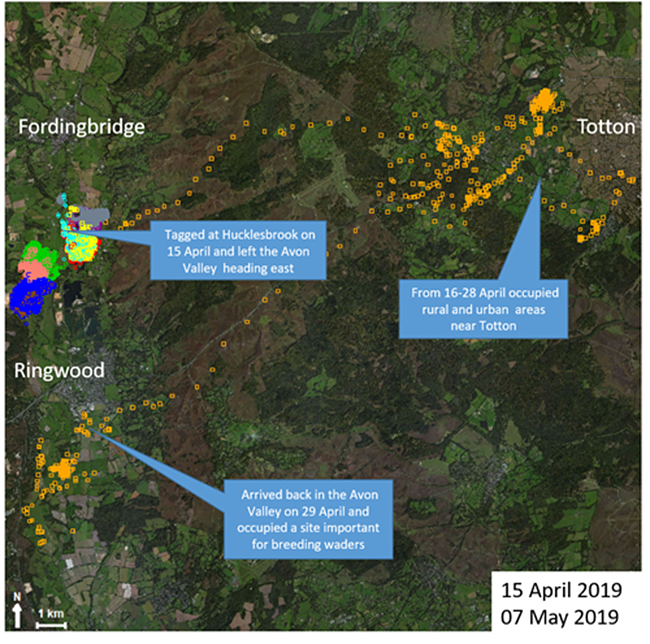
In mid-April 2019 a non-breeding vixen, nicknamed "6644" by the Game & Wildlife Conservation Trust biologist who had fitted her with a GPS collar, was released after having been caught in a snare in a river meadow at Hucklesbrook in Hampshire's Avon Valley. After resting up in a hedge for a few hours she began a purposeful journey eastward across the New Forest, navigating along streams and forestry tracks, to settle 25km (16 miles) away in a rural area near Totton the following day. There she remained for a couple of weeks, moving within about a 4km (2.5 mile) radius. On 28th April, 6644 upped-sticks and took a more or less straight line south-west back to the Avon Valley, arriving on a nature reserve near Ringwood the following day. She moved widely around Ringwood before she was shot by a gamekeeper in early May. The GWCT's GPS data illustrate how some foxes move widely and nomadically, such animals easily covering distances of 20km (12 miles) in a single night.
It has also been established that if a juvenile fails to establish a territory, its activity area becomes smaller and it becomes what German biologist Ad Vos called a "floater" (local itinerant) in a 2003 paper to the Journal of Veterinary Medicine; these animals generally occupy border zones between neighbouring territories.