Deer (Overview) - Taxonomy
There is an expression that goes something along the lines of 'nothing that is worth doing is ever easy', and this certainly seems applicable to the task of classifying mammals. In Linnaeus' time, the situation was a little more straightforward, with animal groups assembled primarily based on the way the critters looked. With the advent of molecular genetics, it has become increasingly apparent that morphology may not always provide the best taxonomic identifiers. Thus, we find ourselves in a situation where different techniques for measuring and weighting taxonomic characters lead to different arrangement of species.
Much has happened in the field of mammal taxonomy in the last half-century and there are many groups, including deer, for which the relationships are still not fully resolved. That which follows is a summary of the situation to date. (Readers interested in more details of how we classify organisms are directed to the Taxonomy page.)
So, let's start with what we can say with any degree of certainty. All of the critters that we know as mammals are grouped together within the class Mammalia. Within this class sits an infraclass (infra being Latin for 'below') called the Eutheria, or "true beasts", which contains all the placental mammals -- i.e., all mammals except monotremes, such as the platypus, and marsupials, like the kangaroos. It is reasonably well established that the Eutheria can be broadly divided into four superorders: the Euarchontoglires (primates, rodents, hares and rabbits); the Xenarthra (anteaters and armadillos); the Afrotheria (elephants and manatees); and, of interest to us here, the Laurasiatheria, which holds the deer along with various other animals including cows, bats and all the carnivores.
Dolphin deer?
The first major sticking point we encounter now is on the placement of the order Cetacea, the whales and dolphins. Molecular data strongly supports the view of renowned mammologist Sir William Flower who, in 1891 proposed that the cetaceans should be grouped with the deer and other related mammals currently placed in the Artiodactyla order. This would form an, as yet unranked, group called the Cetartiodactyla. Despite the molecular data, there is debate among taxonomists as to whether this grouping is a valid taxonomic clade. It seems that while most morphological taxonomists generally support the idea of a clade uniting the two orders, some disagree that the Cetacea evolved from within the Artiodactyla. Instead, they prefer to consider the Cetacea and Artiodactyla sister groups -- i.e., they're more closely related to each other than to any other order. To my mind, the current evidence seems pretty convincing and I suspect further study will validate the Cetartiodactyla.

By this point, you might be wondering what happened to deer being ungulates (i.e., hoofed animals). Back in 1766, Carl von Linne grouped all hoofed mammals under the umbrella (more specifically known as a grandorder) Ungulata, stemming from the Latin unguis, meaning 'nail' or 'hoof'. Of late, however, it has become clear that the ungulates don't represent a genetic unit; that they're not a group of mammals more closely related to one another than to other (non-ungulate) mammals. Rather, it seems that the ungulates form an evolutionary grade. In other words, they're a group whose members have evolved similar adaptations -- hoofs instead of claws, for example. In essence, ungulates still exist as a biological group, just not as a taxonomic one. Indeed, as it currently stands, the Laurasiatheria superorder holds eight orders and the ungulates are split unevenly between two of them: the Artiodactyla and Perissodactyla.
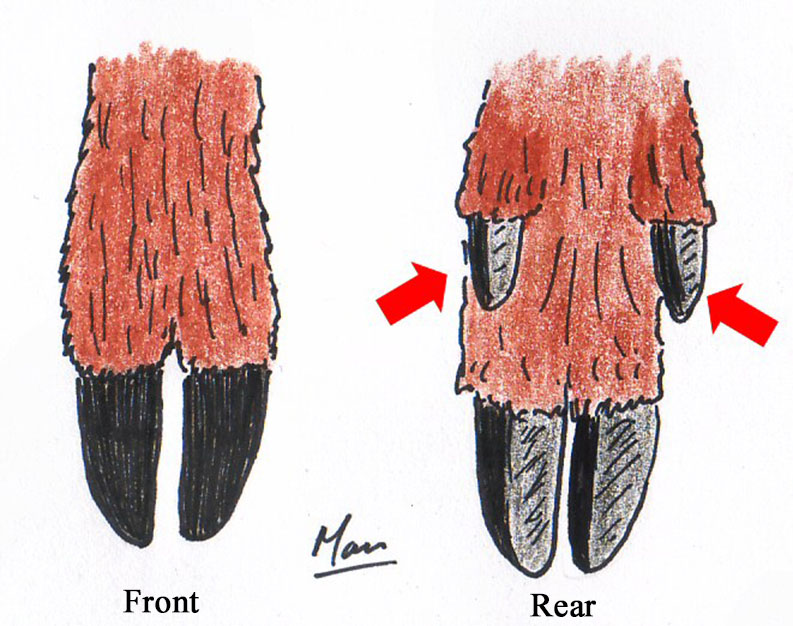
The Perissodactyla, from the Greek perissos meaning 'odd' or 'uneven' and daktulos, meaning 'finger' or 'toe', comprises those mammals with what anatomists call a mesaxonic limb structure; those that walk on the equivalent of the tip of either a single digit (finger/toe) or the tips of three digits, depending on the species. This order contains three families, six genera and about 15 species, including horses, zebras, rhinoceroses and tapirs. Perissodactylian taxonomy is fascinating in its own right, but it is not directly relevant to us here and I won't pursue it further.
The Artiodactyla, from the Greek artios meaning 'even', contains mammals with a paraxonic limb structure; they walk on the third and fourth digits, which are surrounded by keratinizaed hooves. Digits two and five have been reduced to near vestigial status higher up the foot and are called dew claws. In the case of deer and most other ruminants the third and fourth metapodials, the bones in the foot between the ankle and toes, are fused, which makes it seem as though the two toes emerge from the end of a single bone and produces the familiar cloven-hoofed appearance. The order holds ten families, about 80 genera and some 230 species, including pigs, peccaries, hippopotamuses, camels, giraffes, sheep, cows and, of course, deer.
Chewers of the cud
The Artiodactyla can be further divided into four suborders; the one we're interested in is the Ruminatia. It should be mentioned that although all animals within the Ruminatia suborder are ruminants (i.e., they ferment plant matter in their stomachs to aid digestion), not all ruminants are members of the Ruminatia. Llamas and camels, for example, are artiodactyls that ruminate but are part of the Tylopoda suborder rather than the Ruminatia. The Ruminatia can be split into two groups (or infraorders): the Tragulina (the lower ruminants, containing only the chevrotain, or mouse deer) and the Pecora (higher ruminants, containing the "horned" species). Various species sit within the Pecora, including the giraffes and okapis and, of importance here, the Cervidae (the deer family). The Cervidae contains 16 genera and some 51 species. Incidentally, in his Natural History of British Mammals, Derek Yalden suggests that deor was probably the precursor for the modern word "deer", originally it simply meant "beast" and so could be used in reference to any animal.

In a 46 page paper to the Proceedings of the Zoological Society of London during 1878, 19th Century sportsman and naturalist Sir Victor Brooke suggested that the Cervidae could be broadly split into two groups, based on the structure of the bones in their feet. In order to understand this division, we need to take a moment to consider the anatomy of mammalian hands and feet. Your hand is composed of 27 bones (there are 28 in your foot) grouped into three sets. By curling your hand, you can see that your fingers have three sections; these are collectively termed the phalanges, or finger bones and the same name is given to those making up the toes.
Attached to the phalanges at the major knuckle are five longer bones called metacarpals that are connected to a group of smaller bones making up your carpus, or wrist. In the foot, metapodials replace metatarsals and the bones of the heel and ankle are collectively termed the tarsus. It is the metacarpals that are of interest to us here and it is worth noting that anatomists number them from one to five, starting at the thumb or big toe -- so, the thumb's metacarpal is number one, while the pinky's metacarpal is number five.
Brooke split the deer into those that retained only the distil end of the second and fifth metacarpals (i.e., the bits closest to the phalanges/fingers of the index and pinky finger) and those that retained the proximal (carpal/wrist) ends of the same bones; he named them Telemetacarpalian and Plesiometacarpalian deer, respectively. The Telemetacarpalia and Plesiometacarpalia are sometimes also known as the New World or Capreoline and Old World or Cervine species, respectively.

At the time of Brooke's classification, the telemetacarpalian deer included Roe, Chinese water deer, Moose, Black- and White-tailed deer and Reindeer. Plesiometacarpalian deer, included Red, Sika, Muntjac, Fallow, Axis and Pere David's deer. There has been some debate as to the validity of these groups as taxonomic units (which we'll come to in a moment), but most studies have found a split between the deer corresponding roughly to the geographical regions of the Old and New Worlds. Indeed, data from mitochondrial DNA studies suggest that New and Old World deer shared a common ancestor that lived back in the late Miocene, splitting some 9.5 to 12.5 million years ago.
Muddied waters
Since their proposition just over 130 years ago, the validity of the Telemetacarpalia and Plesiometacarpalia as taxonomic units has become a matter of contention. Some geographical and morphological evidence support the division, but many studies have failed to find support for the Telemetacarpalia and, even where support is found, the interrelationships among the species remain unresolved. In their 1987 review of relationships among living deer, for example, Australian Natural University biologist Colin Groves and British-based taxonomist Peter Grubb argued that the lack of antlers in water deer (Hydropotes spp.) is one of the features suggesting that they formed a sister group to all remaining Cervidae species.
If this is the case, the Telemetacarpalia as defined by Brooke isn't a valid taxon. Similarly, in a paper to the Proceedings of the Royal Society of London published during 1998, Ettore Randi at the Istituto Nazionale per la Fauna Selvatica in Italy and colleagues presented cytochrome b data for 11 deer species and failed to find any support for the Telemetacarpalia; they also disagreed with Groves and Grubb, suggesting that Hydropotes is actually nestled with the Black- and White-tailed deer, rather than being the sister group of the Cervidae.

The situation seems less controversial for the Plesiometacarpalia, although recent molecular data has thrown up some interesting placements for deer currently ascribed to the Cervus genus; if borne out, they could invalidate the Plesiometacarpalia. The situation can be rather mindboggling to the non-taxonomist, but the general view seems to be that while the Plesiometacarpalin deer are a valid group, Telemetacarpalin deer aren't. With this in mind, how should the deer be split? Unfortunately, there's no universally accepted answer to this question, but by using a combination of morphological and molecular data, we can hazard a reasonable guess.
Best endevours
The following scheme is based on the major taxonomic analysis of deer (using mitochondrial and nuclear DNA) published in 2006 by a team of French taxonomists at the Museum National d'Historie Naturelle in Paris, led by Clement Gilbert. In their scheme, Gilbert and his team divided the Cervidae family into two sub-families: the Cervinae and the Capreolinae. The deer are then divvied up between the sub-families as follows:
Family: Cervidae (Deer)
Sub-Family: Cervinae (Old World deer)
Tribe: Cervini (True deer)
Genera: Cervus (Red, Sika, Sambar, elk, etc.), Axis (Chital deer), Dama (Fallow deer) and Rucervus (Swamp deer)
Tribe: Muntiacini (Muntjacs)
Genera: Muntiacus (Muntjacs) and Elaphodus (Tufted deer)
Sub-Family: Capreolinae (New World deer)
Tribe: Capreolini (New World deer)
Genera: Capreolus (Roe deer) and Hydropotes (Chinese and Korean water deer)
Tribe: Alceini (Moose)
Genus: Alces (Moose)
Tribe: Odocoileini (Mule deer)
Genera: Rangifer (Reindeer), Odocoileus (Black- and White-tailed deer), Blastocerus (Marsh deer), Pudu (Pudu), Hippocamelus (Andean deer), Mazama (Brocket deer) and Ozotoceros (Pampas deer)
The data presented by Gilbert and his colleagues provide interesting phylogenetic fodder. They suggest that the genus Mazama, currently holding the Brocket deer, may be invalid; so the Brocket deer would need to be reclassified. They assign the Pere David's deer from its own genus (Elaphurus) to the Cervus genus and they move the Barasingha (or Swamp deer) out of Cervus and into its own genus (Rucervus). These amendments don't concern us here, so I won't delve any further into this.

A more recent phylogenetic analysis, published during 2008, in the same journal as Gilbert's study was largely in support of the French biologists' findings, although they made some slight re-arrangements, including considering the tribes as sub-families. The authors also suggested that, until further data are available, the 2006 Cervidae classification proposed by Gilbert and his team should be adopted.
I have followed the 2006 scheme above, to the exclusion of referring to the Cervinae as Plesiometacarpalia and the Capreolinae as Telemetacarpalia, which the 2006 study supported. The data from the 2008 study suggested that the Capreolini and Alceini are more closely related to the Muntiacini and Cervini than to the Odocoileini, which conflicts with the idea of the Telemetacarpalia as a taxon, and I have chosen to follow that here. The 2008 study did find support for Plesiometacarpalia, but I have refrained from using that in the above scheme in a bid to avoid further confusion. Where changes have been made to the grouping of individual genera of deer, I will attempt to provide a summary in the relevant species' section on this site (this includes a treatment of Cervus classification on the Red deer page).
An addendum
There are two final points I would like to clarify before leaving the subject of deer classification.
Firstly, it is worth mentioning that there may be some confusion over the validity of the Odocoilinae as a family. Some sources point out that this family is what's referred to as an "invalid junior synonym" of Capreolinae. In other words, Capreolinae appeared in the literature before Odocoilinae (in 1828 and 1923, respectively) and early trumps recent in taxonomic circles. Some authors, however, still refer to the latter as a valid family (the 2008 scheme referenced earlier, for example). Basically, the problem revolves around the placement of the genera Capreolus, Alces and Hydroptes. If they're included with the remaining American species (i.e., Odocoileus, Hippocamelus, Mazama, etc.) then the correct name is Capreolinae; if they're not, then Odocoilinae is valid.

Secondly, there is another group of deer as distinct from those we've already spoken about: they are the musk deer of the south Asian mountains. These deer appear to be more primitive, in the taxonomic sense, than the Cervidae species; lacking antlers, possessing a gall bladder and a musk gland, the secretion of which is used as a perfume fixative. These seven species are grouped into their own family, the Moschidae, within the Pecora and are the sister group to the Cervidae/Bovidae, so they're more closely related to the "true" deer and the cows, goats and sheep etc., than to the giraffes or the pronghorns that are also in this infraorder. Indeed, several authors have pointed out how Musk deer seem to possess both cervid and bovid features.